

Home
Studies
& Data Analysis
Methods
Microscope studies
Flagella experiment
Laboratory math
Blood fractionation
Gel electrophoresis
Protein gel analysis
Mitochondria
Concepts/ theory
Keeping a lab notebook
Writing research papers
Dimensions & units
Using figures (graphs)
Examples of graphs
Experimental error
Representing error
Applying statistics
Principles of microscopy
Solutions & dilutions
Protein assays
Spectrophotometry
Fractionation & centrifugation
Radioisotopes and detection
Introduction/
training
Mitochondria theory
Mitochondria in vitro
- preparation
- fate of substrates
- state IV
- state III
- metabolic poisons
- mitotraces
- rationale
- experiments
Additional topics
Structure of Mitochondria
The cytoplasm of nearly all eukaryotic cells contain mitochondria, although there is at least one exception, the protist Chaos (Pelomyxa) carolinensis. They are especially abundant in cells and parts of cells that are associated with active processes. For example, in flagellated protozoa or in mammalian sperm, mitochondria are concentrated around the base of the flagellum or flagella. In cardiac muscle, mitochondria surround the contractile elements. Hummingbird flight muscle is one of the richest sources of mitochondria known. Thus, from their distribution alone one would suspect that they are involved in energy production. Multicellular organisms probably could not exist without mitochondria. The inability to remove electrons from the system and the buildup of metabolic end products restrict the utility of anaerobic metabolism. Through oxidative phosphoryation mitochondria make efficient use of nutrient molecules. They are the reason that we need oxygen at all.
The double-membraned mitochondrion can be loosely described as a large wrinkled bag packed inside of a smaller, unwrinkled bag. The two membranes create distinct compartments within the organelle, and are themselves very different in structure and in function.
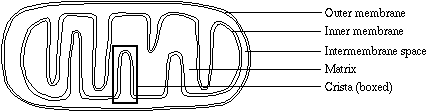
The outer membrane is a relatively simple phospholipid bilayer, containing protein structures called porins which render it permeable to molecules of about 10 kilodaltons or less (the size of the smallest proteins). Ions, nutrient molecules, ATP, ADP, etc. can pass through the outer membrane with ease.
The inner membrane is freely permeable only to oxygen, carbon dioxide, and water. Its structure is highly complex, including all of the complexes of the electron transport system, the ATP synthetase complex, and transport proteins. The wrinkles, or folds, are organized into lamillae (layers), called the cristae (singlular: crista). The cristae greatly increase the total surface area of the inner membrane. The larger surface area makes room for many more of the above-named structures than if the inner membrane were shaped like the outer membrane.
The membranes create two compartments. The intermembrane space, as implied, is the region between the inner and outer membranes. It has an important role in the primary function of mitochondria, which is oxidative phosphorylation.
The matrix contains the enzymes that are responsible for the citric acid cycle reactions. The matrix also contains dissolved oxygen, water, carbon dioxide, the recyclable intermediates that serve as energy shuttles, and much more (see "other functions"). Diffusion is a very slow process. Because of the folds of the cristae, no part of the matrix is far from the inner membrane. Therefore matrix components can diffuse to inner membrane complexes and transport proteins within a relatively short time.
Electron micrographs have revealed the three dimensional structure of mitochondria. However, since micrographs are themselves two dimensional, their interpretation can be misleading. Texts frequently show a picture of a 'typical' mitochondrion as a bacteria-sized ellipsoid (perhaps 0.5 by 1 micrometer). However, they vary widely in shape and size. Electron micrographs seldom show such variation, because they are two-dimensional images.
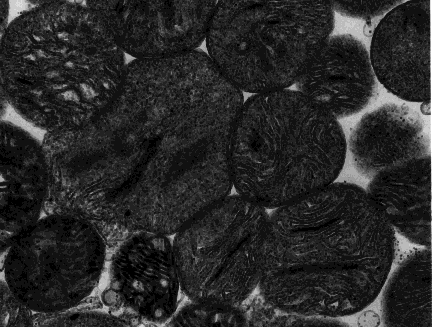
Isolated mitochondria, such as from homogenized muscle tissue, show a rounded appearance in electron micrographs, implying that mitochondria are spherical organelles.

Mitochondria in situ can be free in the cytoplasm or packed in among more rigid structures, such as among the myofibrils of cardiac muscle tissue. In cells such as muscle, it is clear that mitochondria are not spherical, and often are not even ellipsoid. In some tissues, the mitochondria are almost filamentous, a characteristic that two dimensional micrographs may fail to reveal.
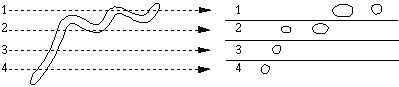
A planar section cuts through one or several parts of the organelle, making a single organelle appear to be more than one. The image we see of a circular or ellipsoidal organelle may disguise the true nature of the mitochondrion.
Visitors: to ensure that your message is not mistaken for SPAM, please include the acronym "Bios211" in the subject line of e-mail communications
Created by David R. Caprette (caprette@rice.edu), Rice University 12 Dec 96
Updated 26 May 05