

Home
Studies
& Data Analysis
Methods
Microscope studies
Flagella experiment
Laboratory math
Blood fractionation
Gel electrophoresis
Protein gel analysis
Mitochondria
Concepts/ theory
Keeping a lab notebook
Writing research papers
Dimensions & units
Using figures (graphs)
Examples of graphs
Experimental error
Representing error
Applying statistics
Principles of microscopy
Solutions & dilutions
Protein assays
Spectrophotometry
Fractionation & centrifugation
Radioisotopes and detection
Guide to the study
Lab part 1
- tutorial/specimens
- Paramecium
- Chlamydomonas
- fixing/observingflagella
- Chaos (Pelomyxa) carolinensis
- Naegleria gruberi
- the five kingdoms
Lab part 2
- experiment introduction
- microtubules
- amputating flagella
- experimental design
- data collection
- class data
Structures and Functions of Microtubules
Microtubules are filamentous intracellular structures that are responsible for various kinds of movements in all eukaryotic cells. Microtubules are involved in nucleic and cell division, organization of intracellular structure, and intracellular transport, as well as ciliary and flagellar motility. Because the functions of microtubules are so critical to the existence of eukaryotic cells (including our own), it is important that we understand their composition, how they are assembled and disassembled, and how their assembly/disassembly and functions are regulated by cells.
For the sake of brevity, only the very basic and universal concepts about microtubules and their organization into flagella will be presented here, leaving many questions unanswered. You will find that textbooks provide more complete descriptions of microtubules and their structures and functions, but they also leave many questions unanswered. Textbooks seldom tell us is how much science knows and does not know about them, and of course they cannot be up to date with the latest discoveries. To fully understand a subject it is important to go to multiple sources. If the subject is especially important to you, you should seek the primary literature, namely original research reports.
"Building blocks" of microtubules - tubulins
All eukaryotic cells produce the protein tubulin, in the usual way. The usual way, of course, is by transcription of genes coding for tubulin to produce messenger RNA, followed by the translation of mRNA by the ribosomes in order to produce protein. Cells maintain at least two types of tubulin, which we call alpha tubulin and beta tubulin. However, it is doubtful that the two types can found in cells as individual proteins.
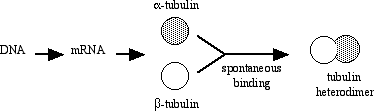
Alpha and beta tubulin spontaneously bind one another to form a functional subunit that we call a heterodimer. A heterodimer is a protein that consists of two different gene products. The term is entirely descriptive - the prefix hetero- means "different," the prefix di- means "two," and the suffix -mer refers to a unit, in this case a single polypeptide. Obviously, cells do not continue to make tubulin (or any other protein) until they run out of resources. Some process must regulate the synthesis of tubulin. A common regulatory mechanism is feedback inhibition.
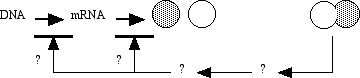
The figure illustrates the inhibition of tubulin synthesis by the presence of heterodimers in the system. Exactly how that inhibition takes place is irrelevant to this discussion. More about the important concept of feedback inhibition can be found elsewhere.
Assembly of microtubules
When intracellular conditions favor assembly, tubulin heterodimers assemble into linear protofilaments. Protofilaments in turn assemble into microtubules. All such assembly is subject to regulation by the cell.
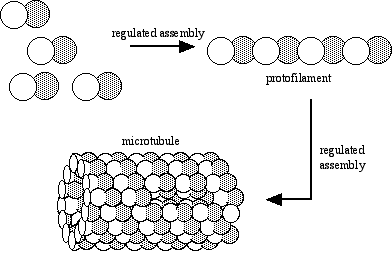
Microtubules form a framework for structures such as the spindle apparatus that appears during cell division, or the whiplike organelles known as cilia and flagella. Cilia and flagella are the most well-studied models for microtubule structure and assembly, and are often used by textbooks to introduce microtubules.
Dynamic instability of microtubules
Under steady state conditions a microtubule may appear to be completely stable, however there is action taking place constantly. Populations of microtubules usually consist of some that are shrinking and some that are growing. A single microtubule can oscillate between growth and shortening phases. During growth, heterodimers are added on to the end of a microtubule, and during shrinkage they come off as intact subunits. The same heterodimer can come off and go back on.
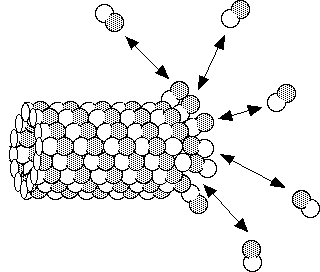
Since even apparently stable microtubular structures have an intrinsic instability, they are considered to be in a dynamic equilibrium, or steady state. Look here to learn about the difference between a steady state and a true equilibrium.
Cilia and Flagella
To understand the regulation of microtubule assembly and function in any organism is a difficult task. To study microtubules in cells as complex vertebrate (e.g., human) cells is a nearly impossible task, without a few "hints" as to how to proceed. The basic mechanisms can be worked out using a much less complex biological model such as a flagellate. For example, the flagella of the photosynthetic protist Chlamydomonas are composed of microtubules, as are all flagella and cilia.
Cilia and flagella have the same basic structure. They are attached to structures known as basal bodies, which in turn are anchored to the cytoplasmic side of the plasma membrane. From the basal bodies the microtubule "backbone" extends, pushing the plasma membrane out with it.
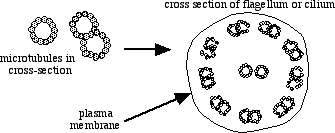
To form cilia or flagella, microtubules arrange themselves in a "9 + 2" array. Each of the two central microtubules consists of a single microtubule with 13 protofilaments arranged to form the wall of a circular tube. Each of the outer nine consists of a pair of microtubules that share a common wall (see the cross sections of microtubules in the figure). Look at the complete cross section carefully. The hair-like appearance of flagella and cilia in a light microscope is misleading. The entire structure lies within the cytoplasm of the cell.
The treatment given here to the structure of microtubules ignores their true complexity. Functional microtubules include associated proteins, anchoring sites and organizing centers, sites for enzyme activity, etc. In cilia and flagella, tubulin forms a core structure to which other proteins contribute structures called dynein arms, radial spokes, and nexin links. The arms, spokes, and links hold microtubules together and allow interaction between microtubules that is superficially similar to the sliding of actin and myosin filaments in muscle contraction.
Ciliary and Flagellar Motion
One might appreciate the complexity of microtubular organelles by looking at the motion of cilia and flagella. Despite the similarities in structure, the difference in nature of motility by flagella versus cilia is profound, as one can see by comparing representatives of the groups Ciliophora (the ciliates) and Mastigophora (the flagellates). Ciliates and flagellates behave differently, live in different habitats and occupy different niches, and likely represent two different evolutionary lineages. The main difference in function is in how they are organized.
Flagella are much longer than cilia and are usually present singly or in pairs. A single flagellum may propel the cell with a whip-like motion. A pair of flagella may move in a synchronized manner to pull the organism through the water, in a way similar to the breast stroke of a human swimmer.
Cilia tend to cover the surface area of a cell. Both cilia and flagella bend as the microtubules slide past one another. The arrangement of cilia permits their coordinated movement in response to signals from the cytoplasm. A small ciliate may have hundreds of individual cilia, all beating in a coordinated manner. How is all of the sliding and bending coordinated? How does the organism "decide" in what direction to move, or how to turn, rotate, or feed? How does it convey the information to hundreds of cilia to bend in a certain way? Questions of that nature are fascinating to cell biologists. They are very difficult to address, because each system is so complex. Nevertheless, with a genome about a hundred times smaller than that of a human, a typical protist is much easier to study than a human cell.
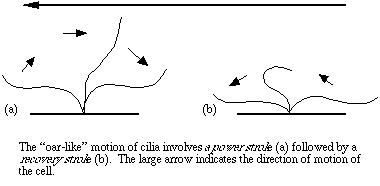
The motion of an individual cilium or flagellum superficially resembles that of an oar, in that it sweeps through the medium with a power stroke that propels the cell. Each power stroke and return stroke involves perhaps thousands of chemical reactions. There may be dozens of strokes per second, and one action may involve thousands of cilia. You may notice that ciliates respond very quickly to obstacles or changes in their environment. It is fascinating to speculate on just how they receive information, process it, and deliver the signals to the cilia to produce precise movement. From the perspective of our relatively slow world, it is also difficult to comprehend how so much can go on in such a short time. Think of the effect on your perception of the universe if you were to shrink by several orders of magnitude.
Visitors: to ensure that your message is not mistaken for SPAM, please include the acronym "Bios211" in the subject line of e-mail communications
Created by David R. Caprette (caprette@rice.edu), Rice University Dates